Permian Reef Complex Virtual Field Trip
Stop I-2: Capitan Reef to Back-Reef Transition
We are a research and service division of:
Carlsbad Caverns National Park entrance sign. Collecting is not permitted in the National Park and permission should be obtained to examine the eastern part of the outcrop which is privately owned. Park vehicles in paved pullof just in front of the National Park entrance sign; cross (carefully!) on foot to south side of road. Cross the dry channel of Walnut Creek and examine spur of rock east of National Park boundary fence, eventually crossing the spur and walking up the next draw to the south (Bat Cave canyon or draw).
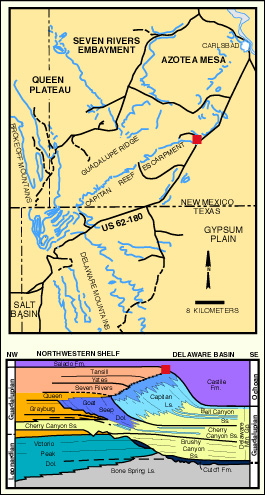
This locality, at the entrance to Walnut Canyon, provides excellent exposures of the reef and near-back-reef facies of the upper Capitan Limestone and Tansill and Yates Formations. In this area, the fore-reef facies and part of the reef have been buried beneath the thick Ochoan (and some thin Tertiary-Quaternary) filling of the Delaware basin. The Castile Fm., although completely or partially removed in areas to the southwest, has been preserved in this area because of the northeastward tilting of this region. Thus, only a small exposure of the reef-crest and its transition to the near-back-reef are exposed. Because an outstanding guidebook is available for the entire Walnut Canyon route (Pray and Esteban, 1977), only rather brief descriptions will be given for these localities (this site corresponds to Locality guide I, stops I and II of Pray and Esteban, 1977).
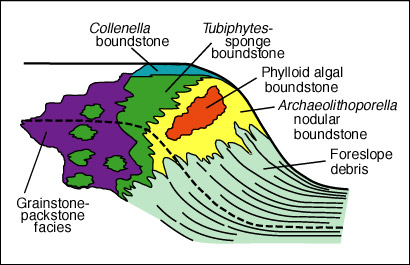
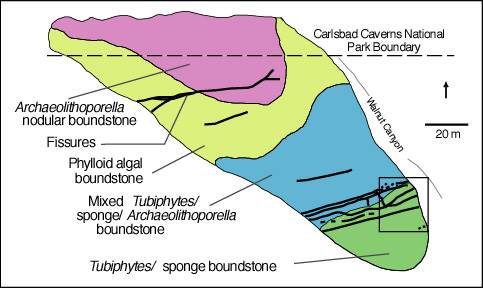
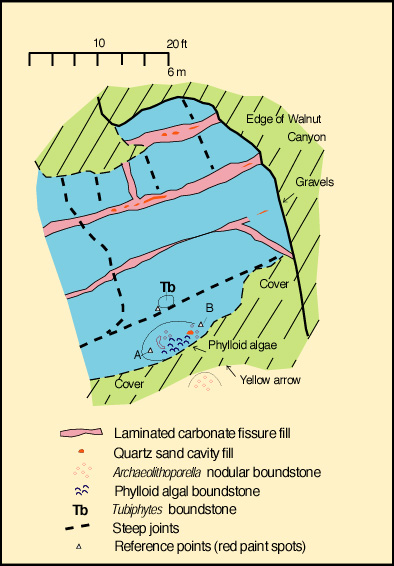
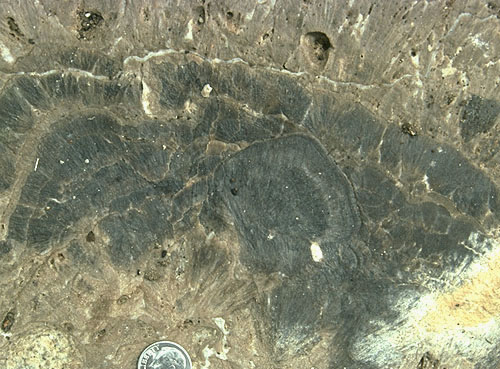
We will examine the rock spur between Walnut Canyon, and Bat Cave Canyon. We will pay particular attention to several fresh outcrops in Bat Cave Canyon for these expose the reef fabric in an unweathered and more readily visible state. Please note, however, that much of this exposure is within the National Park boundaries and thus collecting permits are required for sampling. Babcock (1974) noted a distinct faunal zonation within the reef (see reef microfacies diagram). He recognized an Archaeolithoporella-nodular boundstone, a phylloid algal boundstone, and a Tubiphytes-sponge boundstone/packstone as well as some transitional zones (see map figures below). In all these facies there are four salient elements: 1) an in situ framework of oriented organisms; 2) encrusting and binding organisms which added stability to the framework; 3) internal sediment of skeletal fragments, pellets, or other grains which lodged in open pores in the framework; and 4) submarine cement crusts filling virtually all remnant porosity.
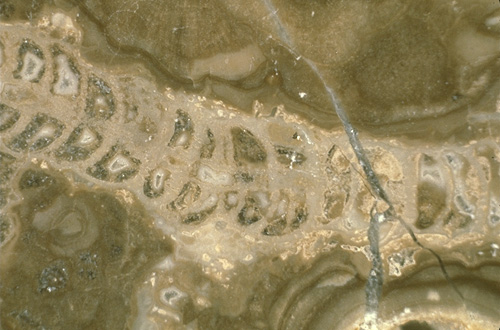
The dominant framework organisms in this complex are the calcareous sponges (see photo). Many different types existed here (see photo), including members of the genera Guadalupia, Amblysyphonella, Cystaulete, and Cystothalamia. Other organisms such as Tubiphytes (of problematic affinities), stromatolitic blue-green algae, phylloid algae (see photo), and bryozoans also form significant framework elements, at least locally. Solitary rugose corals can also be seen locally, although they do not form a volumetrically important part of the reef framework.

© Peter A. Scholle, 1999
© Peter A. Scholle, 1999

© Peter A. Scholle, 1999
Encrustation and stabilization of this skeletal framework was accomplished by Archaeolithoporella (a possible alga), Tubiphytes (found as both framework and encrusting forms), Solenopora (a probable red alga), Collenella (an algal form) and other, less common, organisms (Photo Fig. 18). Such encrustation, seen also in modern reefs, probably contributed greatly to the strengthening of the reef framework.
Internal sediment, although not a significant factor in the lithification of the reef, did play an important role in infilling and occluding the primary reef porosity. Internal sediments generally are found as laminated, geopetal fabrics, sometimes with interlayered submarine cements. The internal sediments contain a specialized fauna including foraminifers, ostracods, echinoids, pelecypods and other organisms which presumably lived within interstices in the reef framework. In areas which are dominated by phylloid algae, internal muddy sediment shows a strongly preferred fabric in the lee side of upright, curved algal blades.
© Peter A. Scholle, 1999
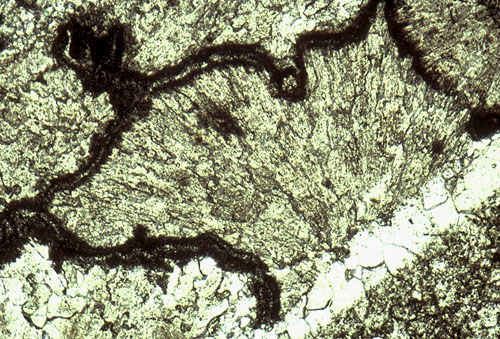
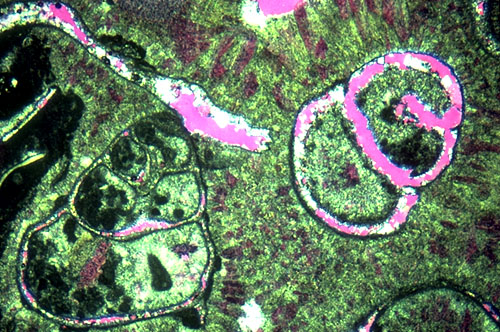
Submarine cements form a very important component of the Permian reef. Coarse fans of radial-fibrous crystals fill much of the primary porosity in the reef and make up more than half the total volume of rock in many locations. The cement fans, probably originally aragonite, are commonly interlayered with Archaeolithoporella or other encrusting organisms (see photo). The submarine cements are restricted to a relatively narrow zone which extends from one to two hundred meters 300-600 ft) down the fore-reef slope to perhaps 0.8 km (1/2 mi) shelfward of the reef crest. Very similar relations have been seen in modern reefs such as in Belize, Florida, the Bahamas, and Jamaica. In these areas, as in the Permian, submarine cementation, largely in the form of aragonitic and high-Mg calcite fans and crusts, are restricted to the reef face, upper fore-reef, and near-back-reef zones (James and Ginsburg, 1979; Macintyre, 1977; Macintyre, 1985). Petrographic evidence has indicated the former existence of cements with both primary aragonite and high-Mg calcite fabrics although both have subsequently been altered to low-Mg calcite (Lohmann and Meyers, 1977; Loucks and Folk, 1976).
Submarine cements are seen filling synsedimentary fractures (see photo) as well as large and small primary cavities in the reef (see photo). The fractures are shelf-margin-parallel features which probably formed due to compaction of underlying reef rubble below a rigid slab of cemented reef. The fractures must have been contemporaneous with sedimentation for they locally are lined with sessile organisms; extensive marine cementation and local silt infiltration into fractures also argues for an early origin. We will see several such fractures at this locality.

© Peter A. Scholle, 1999
© Peter A. Scholle, 1999
After seeing these major fabric elements where they are unweathered and well exposed in Bat Cave Canyon we will cross the spur to Walnut Canyon. Examine the sediments on this traverse and try to recognize the same fabrics where they are more intensely weathered. Also examine the fracture fillings along Walnut Canyon.
© Peter A. Scholle, 1999
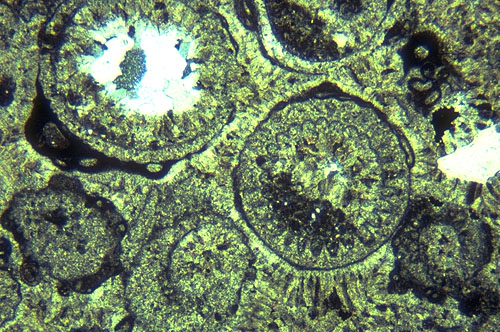
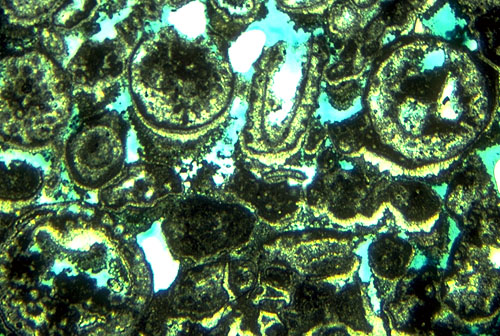
We will now walk up Walnut Canyon examining the transition from reef to near-back-reef areas, eventually crossing from the south to the north side of Walnut Canyon. On this traverse, be sure to note changes in bedding character as well as sediment composition. You should see reef rubble and/or patch reefs. Also note the remarkably rapidity of the lateral lithologic changes. The most obvious change is from a boundstone fabric to one of grainstones and packstones containing ooids and skeletal grains. Cephalopods, foraminifers, pelecypods, gastropods, and most importantly, dasycladacean green algae, particularly Mizzia (see photo) and Macroporella, rapidly supplant sponges and bryozoans as the major skeletal components. Bedding in these well-sorted grainstones is massive and indistinct, but still is far better defined than in the reef facies. Grain size in these near-back reef deposits ranges from coarse silt to coarse sand; sorting is moderately good to excellent, and coated grains and ooids form a significant percentage of the total sediment.
Sediments further up-canyon (farther back- reef) show increasing amounts of dolomite, fenestral fabrics, coated (pisolitic) grains, algal(?) lamination, carbonate breccias, "tepee structures," and thin, clastic terrigenous sand stone-siltstone units.
The abrupt facies transition from reef to back-reef is similar to that seen in many modern settings. In the Florida Keys on the western side of Andros Island in the Bahamas, for example, the change from reefal boundstones to skeletal, back-reef grainstones takes place over distances of just a few tens to hundreds of meters. The near-back-reef areas in Florida and the Bahamas generally consist of complex, small-scale microfacies of green-algal (Halimeda) grainstones, grapestones (coated and coalesced grains), ooids, skeletal fragments, and other lithologies. In areas such as the Joulter's Cay region of the Bahamas, one can see these grainstone types closely intermingled as a series of submarine sand waves, islands, tidal channels, and beaches. Associated with these grainstones are mudstone-wackestone microfacies in sheltered areas of tidal flats and back-barrier coves. The extremely varied lithologies in the Permian near-back-reef setting presumably reflects similarly complex microfacies patterns.
© Peter A. Scholle, 1999
© Peter A. Scholle, 1999
This is also evident in the intimate mixture of diagenetic patterns in the Permian near-back-reef sediments. Submarine, as well as vadose and phreatic nonmarine, cements are all present in local zones in this area, probably as a result of local (island facies) input of nonmarine fluids (see photo of Mizzia green algae above and nearby). Selective leaching of originally aragonitic skeletal fragments, probably in a subaerial setting, led to development of substantial, if localized, secondary porosity (see photo).
A major question in recent years has concerned the nature of the reef margin. Did the reef occupy a position as a topographic high point along the shelf margin or did it form in a slightly deeper water setting on the upper slope (with the ooid-Mizzia grainstones as the shelf edge "high"). Your observations on this traverse should focus, at least in part, on resolving this question.
Return to vehicle(s) and proceed up-canyon.